| 标识符 | CSTR:16397.09.0I01000221 |
|---|---|
| 资源中文名称 | 心肌组织特异性过表达HB-EGF转基因小鼠 |
| 资源英文名称 | B6.Tg(α-MHC-m-HB-EGF)-GC/ILAS |
| 疾病概述 | Hbegf (heparin-binding EGF-like growth factor),HB-EGF信号对于心脏发育和维持是必需的。在心肌肥大或心肌梗死等情况下HB-EGF表达增高。该转基因小鼠可引起心肌纤维化等表型,可用于HB-EGF对心血管系统的生物学功能相关研究。 |
| 实验动物背景信息 | C57BL/6J |
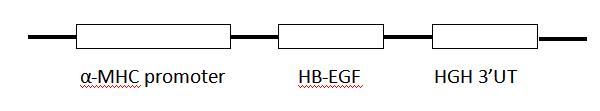
| 模型制作方法 | 扩增并经测序比对正确的HB-EGF片段插入α-MHC启动子下游构建心脏特异表达HB-EGF表达载体。用显微注射法将线性化的转基因载体注射到C57BL/6J小鼠的受精卵中,用ICR小鼠作假孕受体,制备转基因小鼠(图1)。 |
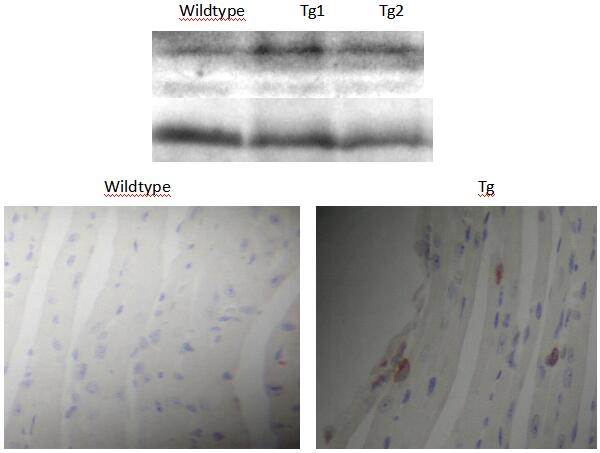
| 模型表型数据 | 1.HB-EGF在转基因小鼠中的高表达和心肌细胞分裂加快(图2)。
2.HB-EGF基因功能 HB-EGF首先在巨噬细胞样U-937细胞的培养介质中被鉴定为EGF家族的成员,是许多细胞类型的有丝分裂原,包括小鼠3T3成纤维细胞,牛主动脉平滑肌细胞,人上皮细胞,小鼠角质细胞,人角质细胞,乳腺和卵巢肿瘤细胞等。HB-EGF通过结合并激活HER-1和HER-4发挥效应。与HER-1结合,细胞发生有丝分裂和趋化反应,而与HER-4结合,仅发生趋化反应。 作为成纤维细胞的有丝分裂原,HB-EGF在皮肤损伤的渗出液中占构成成纤维细胞肝素结合性有丝分裂活性的50%以上,而肝素结合性成纤维细胞有丝分裂原具有刺激成纤维细胞长期增生的潜能。紫外线照射诱导HB-EGF表达,在眼部疾病翼状胬肉中促进上皮细胞和成纤维细胞生长。另外,HB-EGF与肺成纤维细胞增生有关,在钒诱导的肺成纤维细胞增生中HB-EGF表达明显增加,这一过程有MAPK(ERK)信号通路的参与。 肝组织特异HB-EGF转基因小鼠显示HB-EGF过表达可提高部分肝组织切除后肝细胞的增殖,提示HB-EGF可能是肝再生因子。而胰腺组织特异HB-EGF转基因小鼠显示,胰岛间质增多,随年龄增加内分泌部和外分泌部均发生纤维化。这种情况的发生与间质严重扩展有关,提示HB-EGF引起胰岛和间质的相互作用导致胰腺疾病的发生。鉴于成纤维细胞和胰岛的粘附需要EGFR活性,这一信号通路可能参与胰腺疾病的纤维化部分。也许,类似于乳腺组织,胰岛来源的HB-EGF诱导相邻间质细胞产生信号促进导管分支产生。 HB-EGF信号对于心脏发育和维持是必需的。在小鼠胚胎发育晚期,半月瓣和房室瓣边缘表达HB-EGF。HB-EGF基因敲除小鼠显示心室扩张,心脏功能下降,瓣膜明显扩大,提示HB-EGF是一种心脏必需的发育因子。在半月瓣形成过程中,HB-EGF可能调控内皮细胞转化为间质细胞的数量,间质细胞增生的程度,或间质细胞分化为成熟的半月瓣结构,这一过程涉及间质细胞的凋亡。而且HB-EGF部分敲除小鼠显示心内膜HB-EGF表达对心脏正确的发育和行使功能有重要意义。 HB-EGF在心脏中发挥一定的生理功能,而在病理状态下,如心肌肥大或心肌梗死,HB-EGF和/或EGFR家族表达进一步上调。研究发现proHB-EGF分解可导致心肌肥大,而HB-EGF诱导的心肌肥大可能在高血压性心脏病中发挥作用。HB-EGF可能对心脏成纤维细胞的生长发挥调节作用,正如梗死后肾脏实验所提示的一样。体外实验显示HB-EGF mRNA主要表达于培养的成纤维细胞。HB-EGF和EGFR激活的相互作用与心脏成纤维细胞增殖和心肌梗死后心脏重塑相关。 我们的结果显示,野生型小鼠心脏HB-EGF的表达随月龄增加而逐渐降低,而扩张型心肌病转基因小鼠心脏中HB-EGF随月龄增加病情加重而增加,并伴随有间质纤维化以及I型胶原和III型胶原的表达增加。用HB-EGF转基因小鼠研究HB-EGF的功能,发现 HB-EGF可增加心脏间质纤维化,I型胶原和III型胶原的表达。用重组人HB-EGF刺激原代培养心肌成纤维细,30 min即引起I型胶原和III型胶原表达显著增加,24h引起心肌成纤维细胞明显增殖,10 min 即可引起ERK1/2,JNK磷酸化水平增加。 同野生型心脏比较,HB-EGF转基因小鼠左室收缩末期内径(LV systolic diameter)比野生型增加11.5%,左室收缩末期容积(LV systolic volume)增加28.4%。HB-EGF转基因小鼠心肌细胞增殖明显,心脏细胞总数增加。用重组人HB-EGF刺激H9c2细胞10 min 即可引起ERK1/2和JNK磷酸化水平增加。 综上,结果证实HB-EGF参与促进间质纤维化及I型胶原和III型胶原的表达,也促进心肌细胞增殖,导致心脏细胞总数增加。 |
| 动物模型的评价与验证 | 心肌特异表达HB-EGF的转基因小鼠表现为心脏纤维化和猝死等表型,可用于HB-EGF参与心脏纤维化发展以及代偿和失代偿进程等研究。 |
| 保存方式 | 冷冻 |
| 合作方式 | 不限定 |
| 相关文章 |
1.彭健豪, 张连峰. 肝素结合性表皮生长因子(HB-EGF)在心脏发育中的作用[J]. 中国比较医学杂志, 2005,15(4):217-217. 2.彭健豪,邸冉,董伟,张连峰. 肝素结合性表皮生长因子转基因小鼠的建立和功能研究[J]. 中国比较医学杂志, 2007,17(4):187-191. 3.Hong Lian, Yuanwu Ma, Juan Feng, Wei Dong, Qing Yang, Dan Lu, Lianfeng Zhang. Heparin-Binding EGF-like Growth Factor Induces Heart Interstitial Fibrosis via an Akt/mTor/p70s6k Pathway. PLoS ONE. 2012 Sep 12; 7(9): e44946. 4.S. Higashiyama, J.A. Abraham, J. Miller, et al. J.C. Fiddes, M. Klagsbrun, A heparin-binding growth factor secreted by macrophage-like cells that is related to EGF, Science 251 (1991) 936-939. 5.R. Iwamoto, S. Yamazaki, M. Asakura, S. et al. Heparin-binding EGF-like growth factor and ErbB signaling is essential for heart function, Proc. Natl. Acad. Sci. USA 100 (2003) 3221-3226. 6.K. Elenius, S. Paul, G. Allison, J. Sun, et al. M. Klagsbrun, Activation of HER4 by heparin-binding EGF-like growth factor stimulates chemotaxis but not proliferation, EMBO J. 16 (1997) 1268-1278. 7.E. Nishi, A. Prat, V. Hospital, et al. K. Elenius, M. Klagsbrun, N-arginine dibasic convertase is a specific receptor for heparin-binding EGF-like growth factor that mediates cell migration, EMBO J. 20 (2001) 3342-3350. 8.N. Prenzel, E. Zwick, H. Daub, M. et al. EGF receptor transactivation by G-protein-coupled receptors requires metalloproteinase cleavage of proHB-EGF, Nature 402 (1999) 884-888. 9.B. Pan, K. Sengoku, K. Goishi, N. et al. The soluble and membrane-anchored forms of heparin-binding epidermal growth factor-like growth factor appear to play opposing roles in the survival and apoptosis of human luteinized granulosa cells, Mol. Hum. Reprod. 8 (2002) 734-741. 10.D. Nanba, Y. Kinugasa, C. Morimoto, M. Koizumi, H. et al. Loss of HB-EGF in smooth muscle or endothelial cell lineages causes heart malformation, Biochem. Biophys. Res. Commun. 350 (2006) 315-321. |
| 备注 |
 京公网安备 11010502043083号
京公网安备 11010502043083号